CIIL - NEWSLETTER , January 2026 - N° 18
Portraits de Chercheurs

After studying biochemistry in Lyon and gaining teaching experience in Morocco, I completed my PhD at the Biological Chemistry Laboratory at Pierre-et-Marie-Curie University in Paris, under the supervision of Professor R. Acher. I studied structural variations in neurohypophyseal hormones throughout the evolution of vertebrates. This project led to the discovery of a hormone resulting not from a mutation in its gene, but from an alteration in its post-translational maturation. Thus, a single gene could give rise to two hormones with different activities, depending on the enzymatic machinery of the cells expressing it. I then turned my attention to the process of secretion and proteolytic maturation of peptide hormone precursors and joined Dr DF Steiner’s laboratory at the University of Chicago, which had recently identified the endopeptidases responsible for the proteolytic maturation of peptide hormone precursors. I have once again investigated a process of differential proteolytic maturation, namely that of proglucagon, which gives rise to glucagon when expressed in the alpha cells of the islets of Langerhans in the endocrine pancreas, and to glucagon-like peptide 1 (GLP-1) when expressed by L-type enteroendocrine cells of the intestinal epithelium. Interestingly, these two hormones have opposing effects on blood glucose control, and the maturation of their common precursor therefore plays a crucial role in their production. I was able to identify the endopeptidases responsible for this differential maturation and also investigated the mechanisms that regulate the activation of these endopeptidases within the intracellular compartment appropriate to their function.
Following a second postdoctoral fellowship at the Louis-Jeantet Laboratory at the University of Geneva, led by Prof P. Halban, which again focused on the cellular biology of GLP-1, I was recruited by the CNRS and joined the Cell Biology laboratory led by Dr B. Hoflack at the Pasteur Institute in Lille. Having studied the production of peptide hormones during my post-doctoral research, I then turned my attention to the study of a hormone receptor, the leptin receptor, and my scientific interest shifted from the secretory pathway to the endocytic pathway of the cell. It was in this context that I supervised the PhD thesis of Sandrine Belouzard, who is now head of the MCV team, to which I belong.
Following B. Hoflack’s departure, I joined the virology team led by Jean Dubuisson to study viral entry. I naively thought that viruses must enter cells in much the same way as hormones bind to their receptors, but I soon realised that it is more complicated than that. Viral entry is not simply a mechanism of internalisation, but rather a process involving the activation of the virus’s envelope proteins, which enables the virus to fuse its envelope with the cell membrane and thereby introduce its genome into the cytoplasm of the host cell. Some viruses do not even require endocytosis to enter the cell.
Another fascinating aspect of studying viruses is their ability to alter the morphology of intracellular compartments. Many RNA viruses that replicate in the cytoplasm of the host cell possess this ability, which manifests as the creation of a new membrane-bound compartment housing the enzymatic complexes responsible for replicating the viral genome. This occurs through the expression of what is often a very limited number of viral proteins, which act more as command centres than as effectors, recruiting cellular factors to divert them from their physiological functions and make them carry out morphological changes that benefit viral replication. In fact, viruses are far better cellular biologists than we are.
Recently, in the wake of the COVID-19 pandemic, growing awareness of the potential emergence of new dangerous viruses has spurred research into antiviral drugs. In this context, I am developing reporter cell lines to facilitate the screening of antiviral compounds. These cell lines express a fluorescent probe that is localised in the cytoplasm of uninfected cells and moves into the nucleus following infection (see figure). This means that infection can be quantified in live cells without the need for any manual handling, which simplifies the implementation of antiviral compound screens and improves safety for those carrying out these screens, as no additional reagents need to be added to the sealed cell culture plates prior to automated reading of the infection.
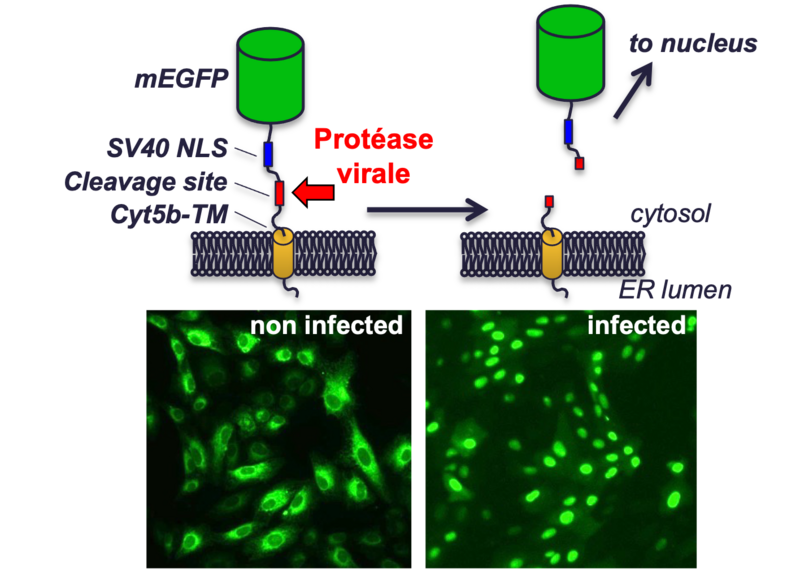
Schematic representation of the fluorescent probe used to detect viral infection. A green fluorescent protein (mEGFP) is anchored to the endoplasmic reticulum membrane via a transmembrane domain (Cyt5b-TM). Between these two domains, two motifs are included: a nuclear localisation site (SV40 NLS) and a proteolytic cleavage site tailored to the specificity of the protease of the virus under study. In uninfected cells, this construct is anchored to the endoplasmic reticulum membrane, which is located in the cytoplasm. When the cell is infected, the viral protease releases the GFP from its membrane anchor and the nuclear localisation site directs it towards the nucleus.

I am a bacteriologist; I arrived in Lille as a postdoc in 1993 to work on Bordetella pertussis in C. Locht’s team. I was appointed to an Inserm CR post in 1995. Throughout my various postings, in Marseille and Lille, I have studied different pathogens: Enterobacter aerogenes (efflux pump), Serratia marcescens (virulence in the C. elegans model), Yersinia pseudotuberculosis and Yersinia pestis (virulence, first in M. Simonet’s team and then in F. Sebbane’s), adherent and invasive E. coli (virulence in the C. elegans model). Having returned to the CIIL in early 2020 to join R. Hartkoorn’s team, I am involved in the development of new antibiotics targeting enterobacteria or Mycobacterium abscessus.


